Sung-Gyu Lee1,
Jong-Bo Kim2,
Hyun Hang1 ![]()
For correspondence:- Hyun Hang Email: hkang@dankook.ac.kr
Received: 18 February 2016 Accepted: 26 May 2016 Published: 28 June 2016
Citation: Lee S, Kim J, Hang H. Anti-oxidative and anti-neuroinflammatory effects of ethyl acetate extract fractions of Suaeda asparagoides MIq. Trop J Pharm Res 2016; 15(6):1175-1181 doi: 10.4314/tjpr.v15i6.9
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the in vitro antioxidant and anti-neuroinflammatory effects of Suaeda asparagoides ethylacetate extract (SAE) in lipopolysaccharide (LPS)-stimulated BV-2 microglial cells.
Methods: The antioxidative activity of SAE was evaluated by measuring 1, 1-diphenyl-2-picryl-hydrazyl (DPPH) radical scavenging activity spectrometrometrically. Cell viability was evaluated by 3-(4, 5-dimethylthiazol-2-yl)-2, 5- diphenyl-tetrazolium bromide (MTT) assay, while LPS-stimulated BV-2 microglia were used to study the ex
Results: Pretreatment with SAE prior to LPS treatment significantly inhibited excessive production of NO (p < 0.001 at 20, 40, 80 and 100 μg/mL) in a dose-dependent manner, and was associated with down-regulation of ex
Conclusion: These results indicate that SAE possesses strong anti-oxidant properties, and inhibits excessive production of pro-inflammatory mediators, including NO, iNOS and TNF-α, in LPS-stimulated BV-2 cells
Introduction
Suaeda asparagoides Miq. (S. asparagoides) is a salt marsh plant which grows well in tide land due to land change [1,2]. Several halophytes are widely distributed along the estuary of Korea [3]. Among them, pharmacological and chemical characterization of Salicornia herbacea have recently been studied well [4]. S. asparagoides plant is traditionally folk medicine in Korea to treat hypertension and hepatitis [4]. Antioxidants are of interest to biologists and clinicians, because they help to protect the human body against damage by reactive free radicals [5,6].
The inflammatory response is mediated by the activated microglia, the resident immune cells of the central nervous system (CNS), which normally respond to neuronal damage [7]. They are activated in response to free radicals and exposure to lipopolysaccharide (LPS) [8]. Activation of microglia results in the release of various bioactive molecules, including nitric oxide (NO), inducible NO synthase (iNOS), interleukins (IL) and tumor necrosis factor (TNF)-α [9]. Overproduction of these inflammatory mediators causes severe neurodegenerative diseases, such as Alzheimer’s disease (AD) and Parkinson’s disease (PD), multiple sclerosis (MS), and Huntington’s disease [9].
LPS is the main component of endotoxins, initiates a number of major cellular effects that play critical roles in the pathogenesis of inflammatory responses, and has been employed to induce microglial activation during infection by Gram-negative bacteria [9]. Therefore, LPS-induced stimulation to microglia might be used as an important tool for evaluating changes caused by activated microglia in vitro. Studies have shown that anti-oxidant and anti-inflammatory agents may inhibit microglial activation and thus protect neuronal cell death seen in various neurodegenerative disorders [10].
Suaeda asparagoides has long been used as a Korean folk herbal medicine for the treatment of functional gastrointestinal disorders [2].However, studies on its beneficial effects on microglia-mediated neuroinflammatory diseases have not been reported. The aim of this study was to investigate whether Suaeda asparagoides ethylacetate extract (SAE) exhibits protective effects on LPS-activated neuroinflammatory processes in BV-2 microglial cells.
Methods
Preparation of S. asparagoides extract
Dried S. asparagoides was purchased from traditional herb market, Seoul, South Korea and was authenticated by a taxonomist Dr. Kim Byung-Wook at Konkuk University, South Korea. A voucher specimen (KH-KU-2015) was kept in our department herbarium for future reference. To obtain the S. asparagoides extract, the dried plant material was ground in a mixer and defatted with three volumes of 70 % ethanol, three times. The residue was extracted with absolute ethanol at 1:10 ratio (w/v) for 2 h in heating mantle at 70 –80 °C. The supernatant was filtered and concentrated in vacuum evaporator system at 50 °C. For further fractionation, alcoholic extract (50 g) was partitioned into hexane, chloroform and ethyl acetate (EA) fractions to furnish 0.67 g, 9.12 g and 10.35 g, respectively. The active EA fraction of S. asparagoides designated as SAE was re-dissolved in distilled water to evaluate for its anti-neuroinflammatory and antioxidant activities.
Evaluation of DPPH radical scavenging activity
The anti-oxidant activity of the SAE was determined using the stable radical 2, 2-diphenyl-1-picrylhydrazyl (DPPH, Sigma-Aldrich, St. Louis, MO, USA). The radical scavenging capacity was evaluated by employing a reaction mixture constituted by aliquots of the SAE and a DPPH methanolic solution as described previously [10]. A sample solution of 60 µL of each SAE, was added to 60 µL of DPPH (60 µM) in methanol. After mixing vigorously for 10 s, the mixture was then transferred into a 100 µL Teflon capillary tube and the scavenging activity of each sample on DPPH radical was measured using a JES-FA ESR spectrometer (Jeol Ltd., Tokyo, Japan). A spin adduct was measured on an ESR spectrometer exactly after 2 min. Experimental conditions were as follows: central field, 3,475 G; modulation frequency, 100 kHz; modulation amplitude, 2 G; microwave power, 5 mW; gain, 6.3 x 105, and temperature, 298 °K. Depending on the sample concentration, ESR spectra were recorded as a function of time starting from the juice and radical solution contact. Recording intervals were 0.5 min and 1.0 min, depending on the sample activity. The signal intensities of radicals were calculated by the double integration of ESR spectra, using the EW (EPRWare) Scientific Software Service program and expressed in arbitrary units.
Cell cultures and determination of cell viability
BV-2 microglia cellswereground in RPMI1640 (Invitrogen, Carlsbad, CA, USA), supplemented with 10 % FBS (Hyclone, Logan, UT, USA) containing 100 U/mL of penicillin(Invitrogen), and 100 μg/mL streptomycin (Invitrogen)at 37 °C in humidified atmosphere of 5 % CO2. In all experiments, cells were pre-treated with the indicated concentration of SAE for 1 h before the addition of LPS (1 µg/mL, Sigma-Aldrich, St. Louis, MO, USA) in serum-free RPMI1640. An equal volume of sterile water was added to all control treatments. Cell viability was determined by 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assay as described previously [10,11]. Cells were incubated with various concentrations of SAE for 24 h followed by MTT for 4 h, and then 100 μL of isopropanol (in 0.04 N-hydrochloric acid) was added to dissolve the formazan crystals. The absorbance was read at 570 nm using the Anthos 2010 spectrophotometer (Salzburg, Austria). Cell viability was calculated as relative absorbance compared to control.
Nitric oxide assay
The amount of NO production in the medium was detected with the Griess reaction [10]. Each supernatant was mixed with the same volume of Griess reagent (1 % sulfanilamide in 5 % phosphoric acid and 0.1 % naphthyl ethylenediamine dihydrochloride in water). The absorbance of the mixture at 540 nm was determined with an ELISA reader (Bio-Tek Instrument, Winooski, VT, USA), nitrite concentration was determined using sodium nitrite as a standard.
Immunoblot analysis
BV-2 cells were lysed in a buffer containing 20 mMTris‐HCl, pH 7.5,150 mMNaCl, 1 mM EDTA, 50 mM β‐glycerophosphate, 1 % NP‐40, 1 mM Na3VO4 and protease inhibitor cocktail (Complete MiniTM, Roche, Mannheim, Germany). The lysates were centrifuged at 15,000 x g for 15 min and supernatants were collected. The concentrations of protein samples were measured using the BCA reagent (Pierce, Rockford, IL, USA). The protein was separated by sodium dodecyl sulphate (SDS) polyacrylamide gel electrophoresis (PAGE) and transferred to PVDF membranes. After blocking with 5 % non‐fat milk, the membranes were hybridized with 1:1000 dilution of primary antibodies for iNOS and β-actin for 12 h at 4 °C. The membranes were washed with Tris‐buffered saline containing 0.1 % Tween‐20 (TBST) three times with 5 min interval and hybridized with horseradish peroxidase‐conjugated mouse or rabbit secondary antibodies for 1 h at room temperature. After washing three times with TBST, antigen‐antibody complexes were visualized with an ECL Western Blot Detection Kit (GE Healthcare) according to the manufacturer’s instructions. The antibodies against iNOS and β-actin were purchased from Cell Signaling Technology INC. (Beverly, MA, USA).
TNF-α assay
BV-2 microglia cells (1 x 105 cells/well) were cultured on 96 well plates and treated with the SAE at indicated concentrations for 1 h and stimulated with LPS (1 µg/mL). At 24 h post LPS treatment, the production of TNF-α was determined in cell supernatant using assay kits (BD Biosciences, San Jose, CA, USA) as per manufacturer’s instructions. The TNF-α assay was performed at room temperature and optical absorbance was measured at 450 nm using ELISA reader.
Statistical analysis
All data are presented as mean ± SEM of at least three independent experiments. Statistical analysis was performed using SAS statistical software (SAS Institute, Cray, NC, USA) by one-way analysis of variance, followed by Dunnett's multiple range tests. P < 0.05 was considered statistically significant.
Results
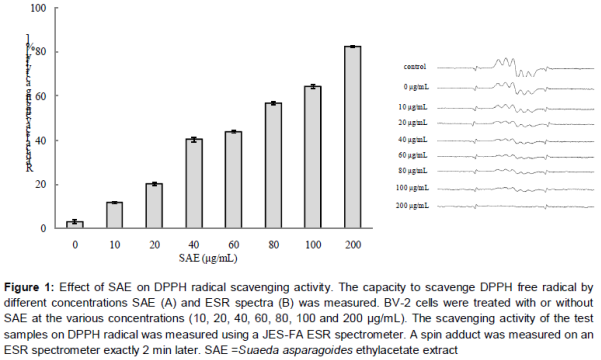
Effect of SAE on DPPH radical scavenging activity
As shown in A, SAE exhibited significant DPPH radical scavenging activity in a dose-dependent manner showing a maximum effect at 200 µg /mL of concentration (p < 0.001). The ESR spectroscopy data at 10, 20, 40, 60, 80, 100 and 200 µg/mL are represented in B.
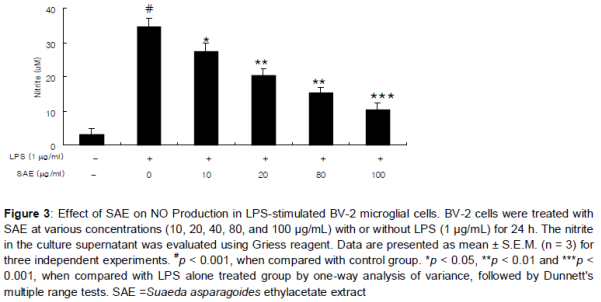
Effect of SAE on LPS-induced NO production
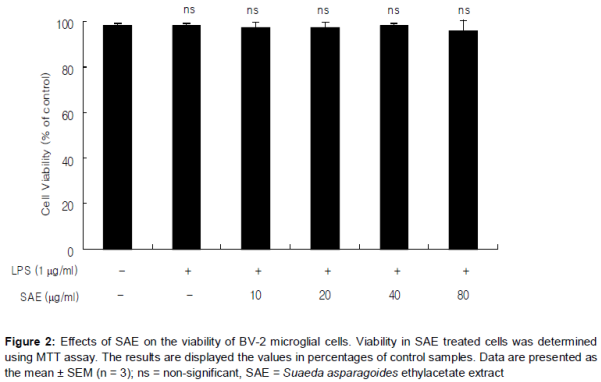
SAE treatment did not exhibit any significant cytotoxicity in BV-2 microglial cells treated for 24 h at concentrations up to 80 µg/mL, and in all cases the viability was found above 95.8 % by MTT assay (). As shown in , treatment of LPS resulted in the excessive production of NO. Pretreatment on SAE significantly suppressed the LPS-induced NO production in a concentration-dependent manner (). The maximum effect was shown at 80 µg/mL (p < 0.001).
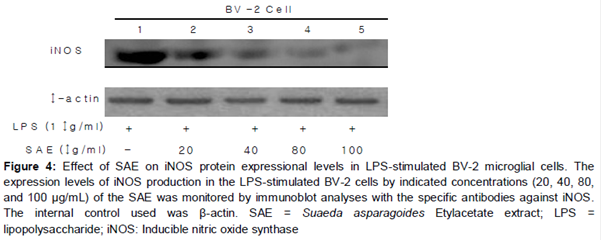
Effect of SAE on LPS-induced expressional levels of iNOS
To know the effect of SAE extract on iNOS levels, BV-2 cells were stimulated with LPS (1 μg/ml) in the presence or absence of SM-EA extract (40, 80 and 100 μg/ml). LPS strongly induced BV-2 cells showing an increased expression of iNOS. However, treatment with SAE exhibited a broad spectrum of inhibitory effect on the expression of iNOS induced by LPS in BV-2 cells ().
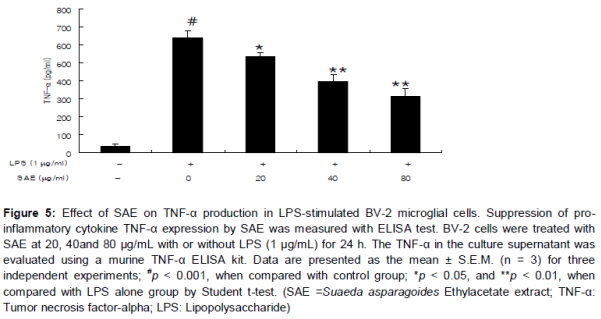
Effect of SAE on TNF-α production in LPS-stimulated BV-2 cells
As shown in , TNF-α levels increased significantly after LPS treatment (1 µg/mL) when compared to those in untreated cells (p < 0.001). However, SAE significantly inhibited TNF-α production in a concentration-dependent manner in LPS-stimulated BV-2 cells (p < 0.05 at 20 µg/mL and p < 0.01 at 40 and 80 µg/mL, respectively).
Discussion
Brown seaweed is a marine organism with various biological activities including anti-inflammatory and anti-oxidative effects [1]. Suaeda asparagoidesis distributed along the shallow marine coast of Korea and is perennially growing on rocks near low tide-level on calm open and sheltered coasts [2]. The present study demonstrated that SAE possesses an antineuro-inflammatory activity in LPS-stimulated BV-2 microglia cells and antioxidant activity.
Furthermore, the SAE exhibited significant antioxidant activity evaluated by DPPH free radical scavenging method. Free radicals and reactive oxygen species (ROS) are important causative factors in the development of age-related neuro-inflammatory and neurodegenerative diseases [12]. Thus neutralization by antioxidants and radical scavengers can reduce neuro-inflammation. It is well known that DPPH radical assay as one of the widely used methods for evaluating the free radical scavenging activities of several antioxidants in a relatively short period of time [13]. In our present study, the SAE also exhibited significant free radical scavenging effect indicating that the SAE might contain antioxidant agents. Since antioxidants might reduce neuro-inflammation [12], we used SAE to evaluate for its anti-neuro-inflammatory activity in LPS-stimulated BV-2 microglial cells.
In BV-2 microglia cells, NO is generated by the inducible isoform of NO synthase (iNOS) and has been described as a neurotoxic substance including nitric oxide [13]. Thus, high levels of NO are produced from L-arginine by iNOS in the brain by prolonged activation of microglial cells, and this response is associated with the progression of various neurodegenerative diseases [14]. Our results clearly show that SAE attenuated LPS-induced iNOS expression and downstream NO production. This result indicates that SAE acts principally by regulating NO generation at the post-transcriptional level and could be beneficial for preventing the progression of neuro-inflammation by BV-2 microglial activation.
Proinflammatory cytokines such as TNF-αare the initiators of the inflammatory response and the mediators of the development of chronic inflammatory diseases [15]. Therefore, the over production of pro-inflammatory cytokines, by activated microglia cell is a possible etiological factor neurological disorders [16]. Microglial cell activation by LPS produces various cytokines including TNF-α leading to the attraction of neutrophils and the accumulation of neutrophil-secreted proteases and ROS at the site of inflammation [13]. SAE may act by suppressing the increased production of TNF-α thereby inhibiting NO production and iNOS expression levels in LPS-stimulated BV-2 cells.
Conclusion
The data demonstrate that SAE has anti-inflammatory properties in LPS-induced BV-2 microglial activation through the down-regulation of inflammation-related gene expression, including iNOS and proinflammatory cytokine such as TNF-α. The strong antioxidant effects exhibited by SAE might also be involved for delivering such potent antineuroinflammatory actions. Therefore, SAE can be considered a useful therapeutic and preventive approach for the treatment of several neurodegenerative diseases and oxidative stress-related diseases.
Declarations
Acknowledgement
References
Archives


News Updates